
Research

I am a biological anthropologist who studies human evolution. My main research interests include the study of hominin (humans and the extinct relatives) taxonomy and phylogeny using cranial morphology, the application of geometric morphometrics to study skeletal variation and functional morphology, and the evolutionary history of Plio-Pleistocene Homo.
I am currently an Associate Professor in the Department of Anatomy at Midwestern University in Glendale, AZ. After earning my Ph.D. in anthropology at the City University of New York (part of the New York Consortium in Evolutionary Primatology) in 2007, I was a Research Instructor in the Department of Anatomical Sciences at Stony Brook University and then an Assistant Professor in the Department of Anthropology at the same university.
Click here for CV (updated 1/2022).
RECENT STUDIES
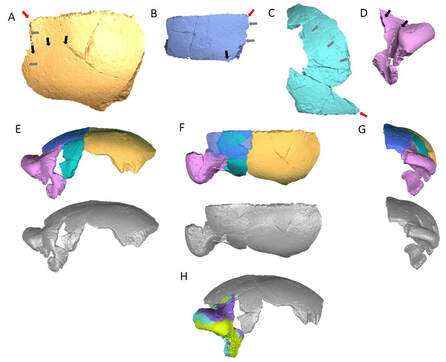
Ongoing Projects
Homo erectus Systematics and Evolutionary History |
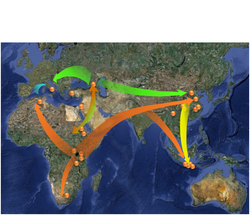
Map of possible Homo erectus sites and hypothetical directions of migration and/or gene flow.
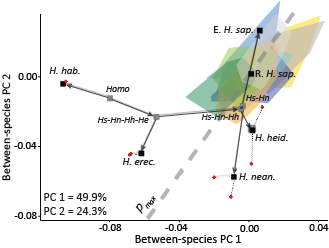
Homo species means and the ancestral node reconstructions plotted in the space with within-population variation. (From Baab, 2018)
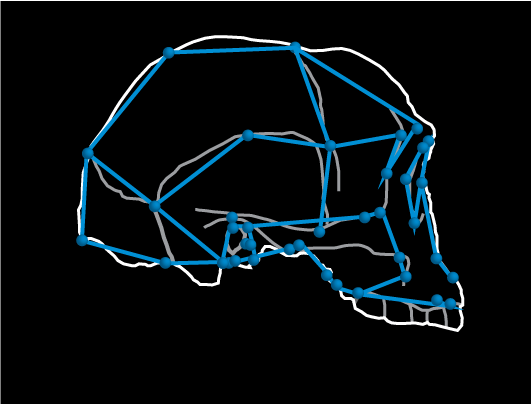
Line drawing of LB1 fossil (type specimen of Homo floresiensis) with 3D landmarks superimposed (blue).
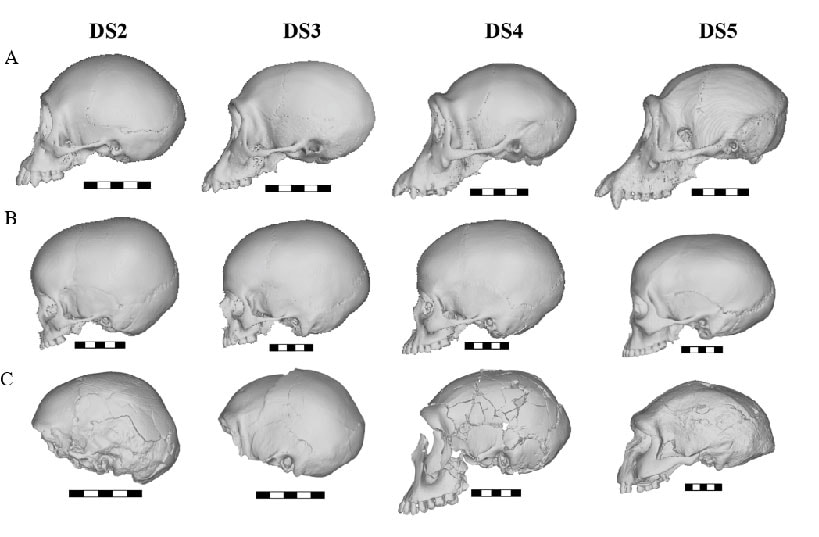
Representatives 3D models of chimpanzees (top), recent humans (middle) and Homo erectus (bottom) from juvenile (left) to adult (right).
|
Recent Peer Reviewed Publications
(most are available on ResearchGate)
Bruner, E., Halloway, R., Baab, K.L., Rogers, M.J., Semaw, S. (2023) The endocast from Dana Aoule North (DAN5/P!): A 1.5 million year-old human braincase from Gona, Afar, Ethiopia. American Journal of Biological Anthropology. 181: 206-215.
Riede, T., Stein, A., Baab, K.L., Hoxworth, J.M. (2023) Post-pubertal developmental trajectories of laryngeal shape and size in humans. Scientific Reports. 13: 7673.
Ward, D. L., Schroeder, L., Roy, J. E., Hertz, M., Uhl, A., Pomeroy, E., Stock, J. T., Copes, L., Baab, K. L., Viola, T. B., Silcox, M. T. (2022) The influence of subsistence strategy and climate on bony labyrinth morphology in recent Homo sapiens. American Journal of Biological Anthropology, 177: 454-470.
Baab, K.L., Rogers, M., Bruner, E., Semaw, S. (2022) Reconstruction and analysis of the DAN5/P1 and BSN12/P1 Gona early Pleistocene Homo fossils. Journal of Human Evolution. 162: 103102.
Baab, K.L., Nesbitt, A., Hublin, J.-J., Neubauer, S. (2021) Assessing the status of the KNM-ER 42700 fossil using Homo erectus neurocranial development. Journal of Human Evolution. 154: 102980.
Baab, K.L. (2021) Reconstructing cranial evolution in an extinct hominin. Proceedings of the Royal Society B. 288: 20202604. Pdf
Riede, T., Coyne, M., Tafoya, B., Baab, K.L. (2020) Postnatal development of the mouse larynx: negative allometry, age-dependent shape changes, morphological integration, and a size-dependent spectral feature. Journal of Speech, Language, and Hearing Research. 63: 2680-94.
Borgard, H.L., Baab, K., Pasch, B. and Riede, T., (2020) The shape of sound: a geometric morphometrics approach to laryngeal functional morphology. Journal of Mammalian Evolution, 27: 577-590.
Baab, K.L. (2018) Evolvability and craniofacial diversification in genus Homo. Evolution. 72: 2781-2791.
Zichello, J., Baab, K.L., McNulty, K.P., Raxworthy, C.J. and Steiper, M.E. (2018) Hominoid intraspcific cranial variation mirrors nuetral genetic diversity. Proceedings of the National Academy of Sciences. 115: 11501-11506.
Baab, K.L., Copes, L.E., Ward, D., Wells, N., Grine, F.E. (2018) Using modern human cortical bone distribution to test the systemic robusticity hypothesis. Journal of Human Evolution. 119: 64-82.
Baab, K.L. and Zaim, Y. (2017) Global and local perspectives on cranial shape variation in Indonesian Homo erectus. Anthrpological Science. 125: 67-83.
Baab, K.L. (2016) The role of neurocranial shape in defining the boundaries of an expanded Homo erectus hypodigm. Journal of Human Evolution. 92: 1-21.
Baab K.L., Brown P., Falk D., Richtsmeier J.T., Hildebolt C.F., Smith K., Jungers W. (2016) A critical evaluation of the Down syndrome diagnosis for LB1, type specimen of Homo floresiensis. PLoS ONE. 11(6): e0155731
Baab, K.L. (2016) The place of Homo floresiensis in human evolution. Journal of Anthropological Sciences. 94: Early View. Link to pdf.
Baab, K.L. (2015) Defining Homo erectus. Henke, W. and Tattersall, I. (Eds.). Handbook of Paleoanthropology. New York: Springer, pp. 2189-2219. *Invited*
Baab, K.L.(2014) Phylogenetic, ecological, and allometric correlates of cranial shape in Malagasy lemuriforms. Evolution. 68: 1450–1468.
Baab, K.L., McNulty, K.P., and Harvati, K. 2013. Homo floresiensis contextualized: A geometric morphometric comparative analysis of fossil and pathological human samples. PLoS ONE. 8(7): e69119.
Baab, K.L., McNulty, K.P., and Rohlf, F.J. 2012. The shape of human evolution: A geometric morphometrics perspective. Evolutionary Anthropology. 21: 151-165.
McNulty, K.P. and Baab, K.L. 2010. Keeping asymmetry in perspective: A reply to Eckhardt and Henneberg. American Journal of Physical Anthropology. 143: 337-339.
Baab, K.L., Freidline, S.E., Wang, S.L., and Hanson, T. 2010. Relationship of cranial robusticity to cranial form, geography and climate in Homo sapiens. American Journal of Physical Anthropology. 141: 97-115.
Baab, K.L. and McNulty, K.P. 2009. Size, shape, and asymmetry in fossil hominins: The status of the LB1 cranium based on 3D morphometric analyses. Journal of Human Evolution. 57: 608-622.
Baab, K.L. 2008. The taxonomic implications of cranial shape variation in Homo erectus. Journal of Human Evolution. 54: 827-847.
Baab, K.L. 2008. A re-evaluation of the taxonomic affinities of the early Homo cranium KNM-ER 42700. Journal of Human Evolution. 55: 741-746.
Other Invited Articles
Delson, E., Baab, K.L. (2011) Fossil Humans in McGraw-Hill Encyclopedia of Science and Technology ed. 10, vol. 7, 464-478. *Invited*
Jungers, W. and Baab, K. 2010. The geometry of hobbits: Homo floresiensis and human evolution. Significance. 6: 159-194.
EVOLUTION SOUP INTERVIEWS
Popular Media Coverage of "Hobbit" Studies
2023: Article on Ancient Origins website Link to article.
2020 YouTube "Evolution Soup" Interview (see video above)
2014 Public Lecture in Erice, Italy:
Interviewed for Italian science website Pikaia. Link to interview (in Italian).
2013:
Featured in the New York Times. Link to article.
Author spotlight blog post on PLoS ONE. Link to blog.
Featured in Le Monde (Paris). Link to article.
Featured in Newsday (Long Island). Download article.
Featured on Planeterde.de. Link to article.
Featured on ATSE Wonder of Science. Link to article.
Featured in Zero Hora (Brazil). Link to article.
2009:
The American Museum of Natural History did a Human Bulletin piece that featured our research and a video is currently featured in the Hall of Human Origins (through Feb. 6, 2009). Navigate directly to the AMNH webpage.
Several on-line news outlets also covered this story, including USA Today, National Geographic, the Guardian (UK), and ScienceDaily. Elizabeth Culotta also mentions the Significance piece in the Science "Origins" blog.
